



Der Nobelpreis für Physiologie oder Medizin wurde den Amerikanern Victor Ambrose von der University of Massachusetts und Gary Ravkan von der Harvard Medical School „für die Entdeckung von microRNA und ihre Rolle bei der posttranskriptionellen Genregulation“ verliehen.
Die in den Chromosomen gespeicherten Informationen können mit Anweisungen für alle Zellen im Körper verglichen werden. Jede Zelle enthält die gleichen Gene und die gleichen Anweisungen. Allerdings unterscheiden sich verschiedene Zelltypen, insbesondere Muskel- und Nervenzellen, erheblich voneinander und erfüllen unterschiedliche Funktionen. Diese Unterschiede werden durch eine mehrstufige Genregulation gewährleistet, wodurch in jedem Zelltyp ein anderer Satz von Genen aktiv ist und die notwendigen Anweisungen angewendet werden. Die Preisträger entdeckten einen der Mechanismen der Genregulation, bei dem sehr kurze RNA-Moleküle, sogenannte microRNAs, eine Schlüsselrolle spielen. Je komplexer der Organismus, desto mehr Arten von microRNAs enthält er: Etwa 2000 davon wurden beispielsweise bereits beim Menschen entdeckt. Die Entdeckung der Preisträger wurde bisher nicht in der Medizin genutzt, könnte aber bei der Behandlung von Krebs nützlich sein .
Obwohl microRNAs in einem kleinen Teil des Genoms kodiert sind, erfüllen sie unter normalen physiologischen Bedingungen eine Reihe von Schlüsselfunktionen bei der Entwicklung, Zelldifferenzierung, Regulierung der Genexpression, dem Zellzyklus und der Apoptose, schreibt die Zeitschrift Biomolecule.
Diese kleinen Regulatoren sind ein äußerst wichtiges Werkzeug für den Körper. Die Störung einer oder einer kleinen Untergruppe von microRNAs hat tiefgreifende Auswirkungen auf die Expressionsmuster von Hunderten von mRNAs. Und dies wiederum führt wie in einer Kette zu Störungen im Ablauf grundlegender biologischer Prozesse der Zelle. Das Ergebnis all dieser Veränderungen ist häufig ein Tumor. So verwandelten sich microRNAs im Handumdrehen von wichtigen Controllern des Zellzyklus in Saboteure für ihren eigenen Körper. Dieses Verhalten ist typisch für onkogene microRNAs.
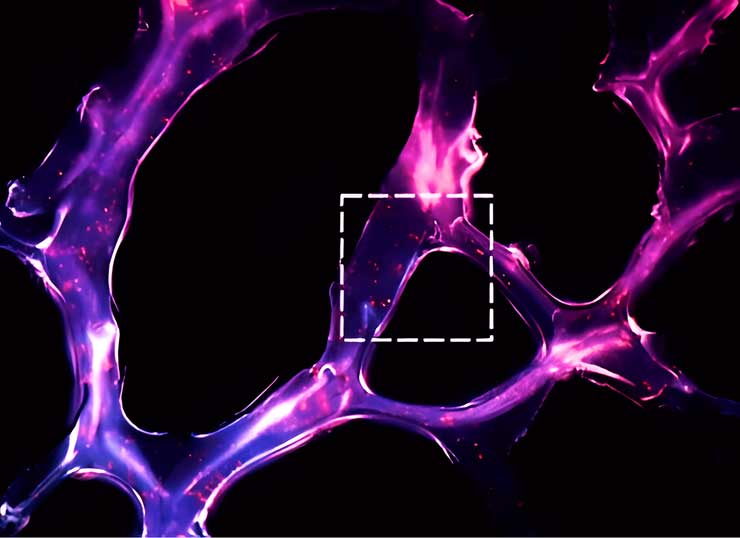
Selbstorganisiertes RNA-Triple-Helix-Hydrogel-Gerüst für die microRNA-Modulation in der Tumormikroumgebung. Kryoschnitt eines Dendrimer-Dextran-Klebstoffhydrogels (12 µm dick), der die Morphologie des Klebstoffs zeigt (Dextranaldehyd wurde mit Alexa Fluor 405 markiert). Rote Punkte stellen Triple-Helix-Nanopartikel dar, die die Oligonukleotide Q705 (rot) und Q570 (grün) enthalten.
Es gibt aber auch Tumorsuppressor-microRNAs, die, als ob sie versuchen würden, sich selbst zu rehabilitieren, Tumorbildungsprozesse hemmen und die Krebsremission fördern. Diese Einteilung ist sehr unklar, da einige microRNAs unter bestimmten Bedingungen tumorsuppressiv und unter anderen onkogen wirken können.
Basierend auf solch gegensätzlichen Funktionen ist es schwierig, eindeutige Schlussfolgerungen über die Rolle von microRNAs bei der Krebsentstehung zu ziehen, aber zahlreiche Studien haben ihren Einfluss auf den Zellzyklus, die Metastasierung und die Angiogenese im Zusammenhang mit der Krebsentstehung detailliert beschrieben.
Nicht jede Krankheit kann sich einer solchen Resonanz in der Gesellschaft rühmen wie Krebs. Dieser Sammelbegriff umfasst eine große Gruppe von Krankheiten, die ohne Übertreibung als „globales Problem“ bezeichnet werden können. Das Thema Krebsbehandlung bleibt bis heute aktuell. Nach Angaben der WHO forderte die Krankheit im Jahr 2020 etwa 10 Millionen Todesopfer, was etwa einem Sechstel aller Todesfälle entspricht.
MicroRNAs (miRNAs) sind eine Klasse kleiner nichtkodierender RNA-Moleküle, deren Sequenzen durchschnittlich 20 Nukleotide lang sind. Sie werden normalerweise im Kernel zusammengestellt und folgen dabei zwei Hauptpfaden: kanonisch und nicht-kanonisch.
Die erste ist mit der Transkription primärer microRNA (pri-microRNA, vom englischen „primary“ – „primär“) unter Verwendung der RNA-Polymerasen II oder III verbunden. Diese pri-miRNA wird durch eine Ribonuklease namens Drosha zu einem microRNA-Vorläufer (prä-miRNA) verarbeitet. Das Endergebnis wird vom Zellkern in das Zytosol exportiert, wo eine Endonuklease der RNase III-Familie (Dicer) die prä-miRNA in einen miRNA-Duplex umwandelt (Abbildung 1).
Die Reifung von microRNAs endet mit der Bildung eines Ribonukleoproteinkomplexes, miRISC (microRNA-induzierter Silencing-Komplex). Im Zytoplasma assoziieren microRNAs mit den Proteinen TRBP (Transactivation Response RNA-Binding Protein), PACT (Protein Kinase R Activating Protein) und einem der Proteine der Argonaute-Familie, wodurch der RISC-Komplex entsteht. Erst nach der Bildung dieses Komplexes erlangt microRNA funktionelle Aktivität. Der andere microRNA-Strang (Passagier) wird oft (aber nicht immer) entfernt.
Nach dem beschriebenen Mechanismus erfolgt die Biogenese der meisten microRNAs, aber dieser Weg ist nicht der einzige. Etwa 40 % der tierischen miRNAs werden in Introns proteinkodierender Gene kodiert und daher sind viele pri-miRNA-Transkripte sowohl prä-mRNA als auch pri-miRNA (Cai et al., 2004; Kim et al., 2009). Solche Introns, Mirtrons genannt, werden gespleißt, um miRNA-Haarnadeln zu bilden, die für die Dicer-Spaltung geeignet sind. In diesem Fall umgeht die microRNA einige Stufen des kanonischen Signalwegs.
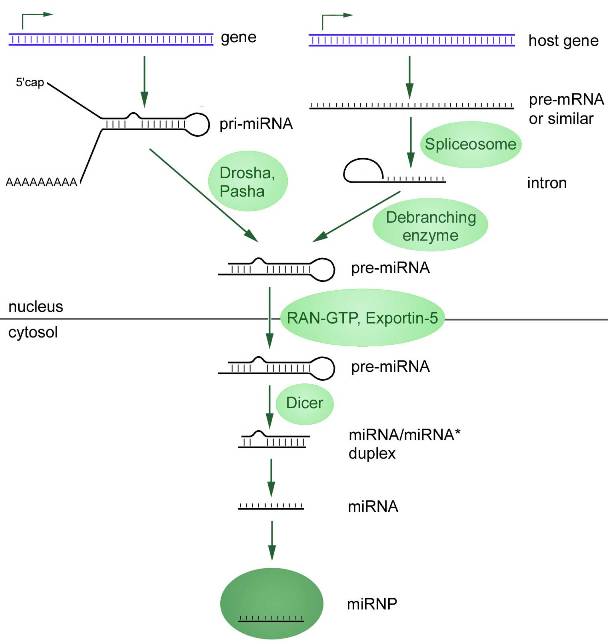
Abbildung 1. Zwei Wege der microRNA-Biogenese, beginnend im Zellkern mit der Transkription. Der kanonische Weg ist links dargestellt: pri-microRNA unterliegt einer 5′-Verkappung und häufig einer 3′-Polyadenylierung (mit Ausnahme von Genen neben Alu-Wiederholungen). Mit Hilfe von Drosha Pasha-Enzymen wird die pri-microRNA beschnitten und in eine Vorläufer-miRNA (prä-microRNA) mit kürzerer Sequenz umgewandelt. Nicht-kanonischer Weg: Primäre microRNA (pri-microRNA) wird von Introns (Mirtrons) im Zellkern transkribiert, die durch Spleißosomen weiter gespleißt werden und dann eine charakteristische Haarnadel bilden. Die durch zwei Synthesewege gebildete Prä-microRNA wird vom Zellkern in das Zytoplasma exportiert, wo sie durch das Dicer-Enzym in reife microRNA umgewandelt wird.
Es gibt viele weitere Varianten der microRNA-Biogenese, aber diese beiden können als die wichtigsten bezeichnet werden.
Die Rolle von microRNAs sollte nicht unterschätzt werden. Dies sind sehr wichtige Regulatoren vieler biologischer Prozesse. Die Tatsache, dass etwa ein Drittel der Protein-kodierenden Gene durch microRNAs gesteuert werden, zeigt deutlich den weitreichenden Einfluss dieser Moleküle, der direkt oder indirekt nahezu alle zellulären Signalwege beeinflusst. Der Hauptfaktor für die Wirkung von microRNAs ist die Regulierung der Genexpression durch Unterdrückung, Blockierung ihrer Translation oder Zerstörung spezifischer Boten-RNA-Transkripte (mRNA).
Die regulatorische Aktivität von microRNAs steht in engem Zusammenhang mit dem RISC-Proteinkomplex, der für zwei wichtige Schritte verantwortlich ist: die Zielerkennung und die Umsetzung des regulatorischen Effekts. Die Erkennung der Ziel-mRNA und die Bindung werden durch das komplementäre Zusammenspiel der microRNA-Erkennungsstelle und der in der Ziel-mRNA lokalisierten Erkennungsregion (miRNA-Response-Element, MRE) sichergestellt. Die regulatorischen Wirkungen von microRNAs werden durch viele Proteinkomponenten von RISC gewährleistet, wie z. B. AGO (Argonaute), das nach Erkennung und Bindung an das Ziel den Prozess der Entwicklung der Wirkung auf das Ziel auslöst.
Der RISC-Proteinkomplex ist eine sehr wichtige Struktur, die an vielen Prozessen zur Regulierung der Genaktivität beteiligt ist (Abbildung 2A).
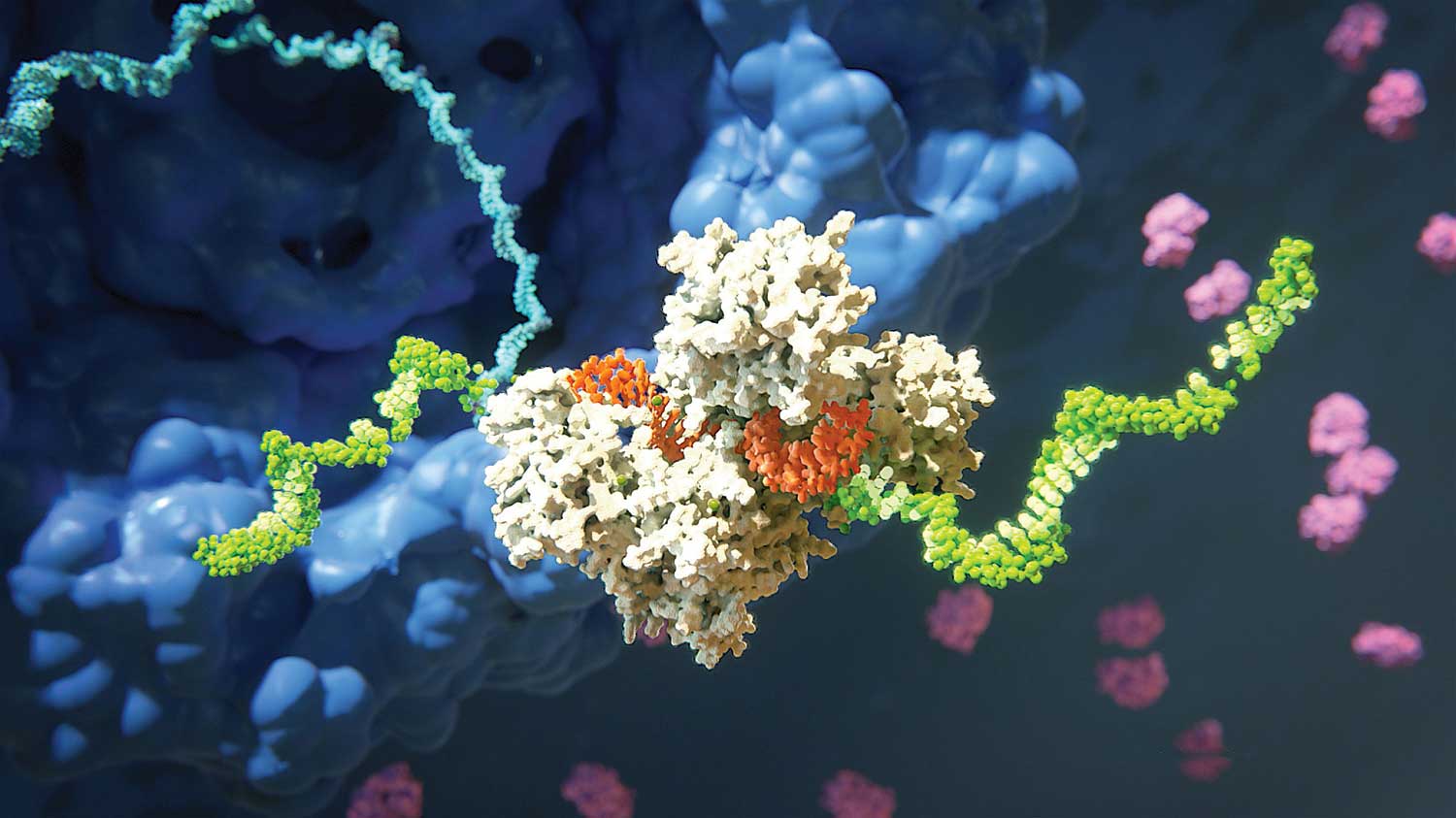
Abbildung 2A. RNA-induzierter Komplex (RISC), der Ziel-mRNA spaltet. RISC (hier weiß dargestellt) umfasst das Argonaute-Protein und ein einzelsträngiges RNA-Molekül (orange), das von Small Interfering RNA (siRNA) abgeleitet ist. Es kommt zur Spaltung der mRNA (grün), die zu einer der microRNA-Ketten komplementär ist. Dieser Prozess endet mit dem Abbau der Ziel-mRNA.
Die Interaktion von microRNA mit dem RISC-Proteinkomplex erfolgt nach folgendem Szenario: Einer der Stränge des miRNA-Duplex (führend) wird in den Proteinkomplex eingefügt und bildet eine Struktur, die das 3′-untranslatierte Ende (3′-UTR) von bindet die Ziel-mRNA. Danach wirkt die microRNA auf ihr Ziel ein, was zur Zerstörung der mRNA durch Nukleasen oder zur Unterdrückung ihrer Translation führen kann (Abbildung 2B).
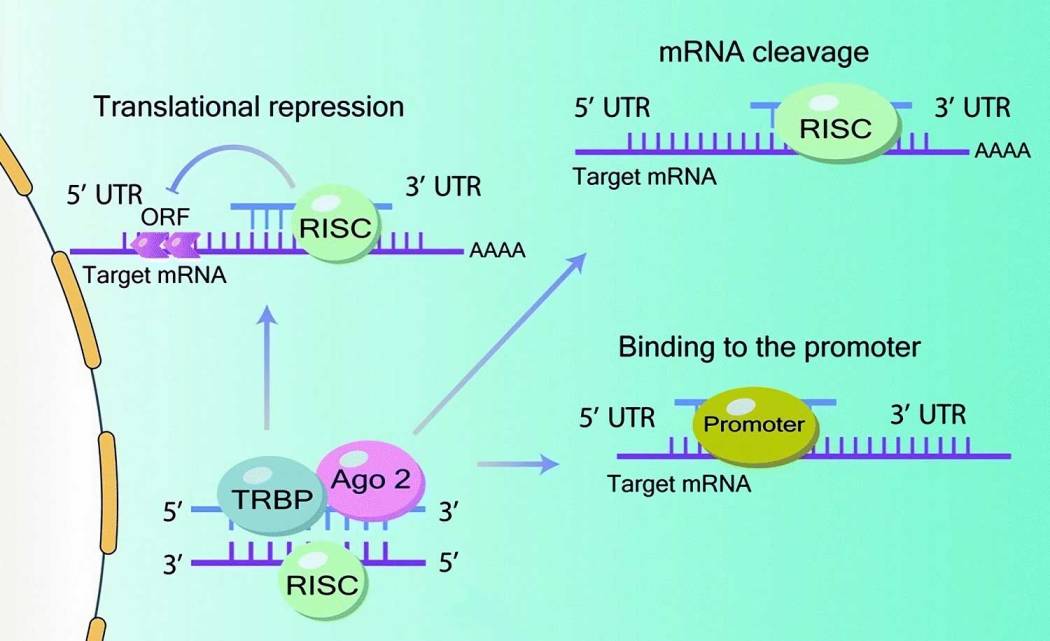
Abbildung 2B. Wirkungsmechanismus von microRNA: Nach dem Export von prä-microRNA aus dem Zellkern (unter Verwendung von Ran-GTP-abhängigem Exportin-5) in das Zytoplasma und der Umwandlung in reife microRNA erfolgt der Aufbau des RISC-Komplexes durch die Interaktion der RNA- Bindungsprotein TAR (TRBP) und Argonaute 2 (Ago 2), um auf die 3′-untranslatierten Regionen (3′-UTR) der Ziel-mRNAs abzuzielen. Der Translationsprozess wird dann durch komplementäre Bindung an das 3′-untranslatierte Ende (3′-UTR) der Ziel-mRNA gehemmt. Es sind jedoch auch andere Arten von Wechselwirkungen möglich: Abbau der mRNA oder Hemmung der Translation aufgrund der Bindung an den Promotor der kodierenden Region und der 5′-UTR.
Im Allgemeinen geht man davon aus, dass tierische miRNAs überwiegend durch translationale Unterdrückung funktionieren, wohingegen pflanzliche miRNAs durch posttranskriptionelle Unterdrückung der Genexpression funktionieren. Gleichzeitig sind microRNAs in der Lage, die Translation in den Phasen der Initiation und Elongation zu beeinflussen. Interessanterweise wurden neben der Transkriptionsunterdrückung auch andere Wirkmechanismen von microRNAs entdeckt. Somit können microRNAs mRNAs stabilisieren und ihre Translation verbessern, als „Köder“ fungieren, die Interaktion von Proteinfaktoren mit ihren RNA-Zielen verhindern und auch an der Reifung anderer microRNAs beteiligt sein.