



El Premio Nobel de Fisiología o Medicina fue otorgado a los estadounidenses Victor Ambrose de la Universidad de Massachusetts y Gary Ravkan de la Facultad de Medicina de Harvard “por el descubrimiento del microARN y su papel en la regulación genética postranscripcional”.
La información almacenada en los cromosomas se puede comparar con las instrucciones de todas las células del cuerpo. Cada célula contiene el mismo conjunto de genes y el mismo conjunto de instrucciones. Sin embargo, los diferentes tipos de células, particularmente las musculares y nerviosas, son bastante diferentes entre sí y realizan funciones diferentes. Estas diferencias están garantizadas por la regulación genética de múltiples niveles, por lo que en cada tipo de célula está activo un conjunto diferente de genes y se aplican las instrucciones necesarias. Los galardonados descubrieron uno de los mecanismos de regulación genética, en el que moléculas de ARN muy cortas llamadas microARN desempeñan un papel clave. Cuanto más complejo es el organismo, más variedades de microARN contiene: por ejemplo, ya se han descubierto alrededor de 2.000 en humanos. El descubrimiento de los galardonados aún no se ha utilizado en medicina, pero puede ser útil en el tratamiento del cáncer. .
Aunque los microARN están codificados en una pequeña parte del genoma, en condiciones fisiológicas normales desempeñan una serie de funciones clave en el desarrollo, la diferenciación celular, la regulación de la expresión genética, el ciclo celular y la apoptosis, escribe la revista Biomolecule.
Estos pequeños reguladores son una herramienta sumamente importante para el organismo. La alteración de uno o un pequeño subconjunto de microARN tiene efectos profundos en los patrones de expresión de cientos de ARNm. Y esto, a su vez, como en cadena, conlleva alteraciones en el curso de los procesos biológicos fundamentales de la célula. El resultado de todos estos cambios suele ser un tumor. Entonces, en un instante, los microARN pasaron de ser importantes controladores del ciclo celular a saboteadores de su propio cuerpo. Este comportamiento es típico de los microARN oncogénicos.
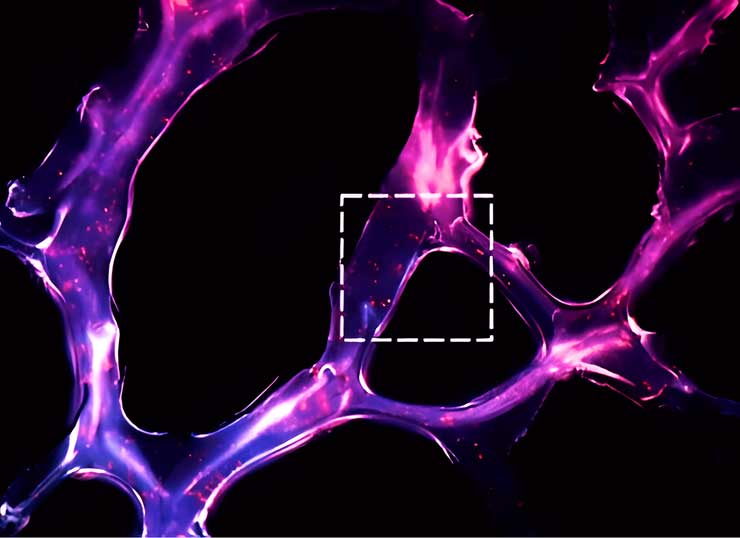
Andamio de hidrogel de triple hélice de ARN autoensamblado para la modulación de microARN en el microambiente tumoral. Criosección de un hidrogel adhesivo de dendrímero-dextrano (12 µm de espesor) que muestra la morfología del adhesivo (el aldehído de dextrano se marcó con Alexa Fluor 405). Los puntos rojos representan nanopartículas de triple hélice que contienen los oligonucleótidos Q705 (rojo) y Q570 (verde).
Pero también existen microARN supresores de tumores que, como si intentaran rehabilitarse, inhiben los procesos de formación de tumores y favorecen la remisión del cáncer. Esta división es muy ambigua, ya que algunos microARN pueden ser supresores de tumores en algunas condiciones y oncogénicos en otras.
Basándose en funciones tan opuestas, es difícil sacar conclusiones claras sobre el papel de los microARN en la carcinogénesis, pero numerosos estudios han detallado su influencia en el ciclo celular, la metástasis y la angiogénesis en el contexto del desarrollo del cáncer.
No todas las enfermedades pueden presumir de tanta resonancia en la sociedad como el cáncer. Este término colectivo abarca un amplio grupo de enfermedades que, sin exagerar, pueden denominarse “problema global”. La cuestión del tratamiento del cáncer sigue siendo relevante hasta el día de hoy. Según la OMS, la enfermedad se cobró alrededor de 10 millones de vidas en 2020, lo que equivale aproximadamente a una sexta parte de todas las muertes.
Los microARN (miARN) son una clase de pequeñas moléculas de ARN no codificantes, secuencias con una longitud promedio de 20 nucleótidos. Generalmente se ensamblan en el kernel, siguiendo dos caminos principales: canónico y no canónico.
El primero está asociado con la transcripción de microARN primario (pri-microARN, del inglés primario – “primario”) utilizando ARN polimerasas II o III. Este pri-miARN se procesa en un precursor de microARN (pre-miARN) mediante una ribonucleasa llamada Drosha. El resultado final se exporta desde el núcleo al citosol, donde una endonucleasa de la familia RNasa III (Dicer) convierte el pre-miARN en un dúplex de miARN (Figura 1).
La maduración de los microARN finaliza con la formación de un complejo de ribonucleoproteína, miRISC (complejo silenciador inducido por microARN). En el citoplasma, los microARN se asocian con las proteínas TRBP (proteína de unión a ARN de respuesta a la transactivación), PACT (proteína activadora de la proteína quinasa R) y una de las proteínas de la familia Argonaute, como resultado de lo cual se forma el complejo RISC. Sólo después de la formación de este complejo el microARN adquiere actividad funcional. La otra cadena de microARN (pasajera) a menudo (pero no siempre) se elimina.
Según el mecanismo descrito se produce la biogénesis de la mayoría de los microARN, pero esta vía no es la única. Aproximadamente el 40% de los miARN animales están codificados en intrones de genes codificadores de proteínas y, por lo tanto, muchas transcripciones de pri-miARN son tanto pre-mARN como pri-miARN (Cai et al., 2004; Kim et al., 2009). Estos intrones, llamados mirtrones, se empalman para formar horquillas de miARN adecuadas para la escisión de Dicer. En este caso, el microARN pasa por alto algunas etapas de la vía canónica.
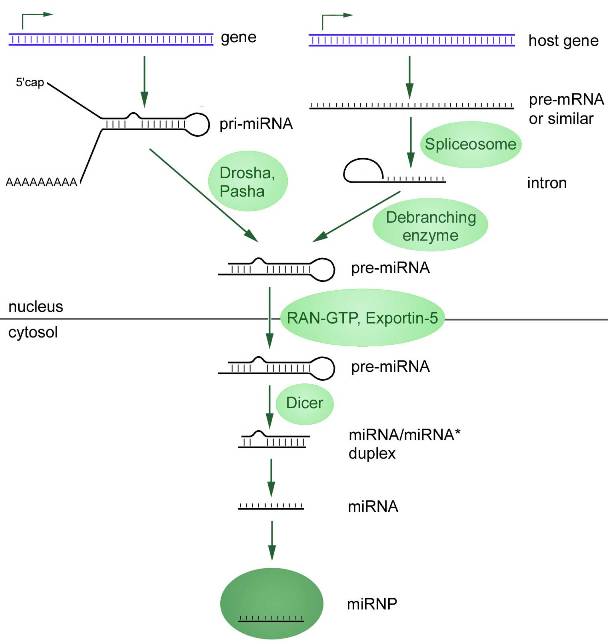
Figura 1. Dos vías de biogénesis de microARN, comenzando en el núcleo celular con la transcripción. La vía canónica se presenta a la izquierda: el pri-microARN sufre una protección en 5 ‘y, a menudo, una poliadenilación en 3’ (la excepción son los genes adyacentes a las repeticiones de Alu). Con la ayuda de las enzimas Drosha Pasha, el pri-microARN se recorta, transformándose en un miARN precursor (pre-microARN) con una secuencia más corta. Vía no canónica: el microARN primario (pri-microARN) se transcribe a partir de intrones (mirtrones) en el núcleo, que luego se empalma mediante espliceosomas y luego forma una horquilla característica. El pre-microARN formado como resultado de dos vías de síntesis se exporta desde el núcleo al citoplasma, donde se convierte en microARN maduro a través de la enzima Dicer.
Hay muchas más variantes diferentes de la biogénesis de microARN, pero estas dos pueden considerarse las principales.
No se debe subestimar el papel de los microARN. Estos son reguladores muy importantes de muchos procesos biológicos. El hecho de que aproximadamente un tercio de los genes que codifican proteínas estén controlados por microARN indica claramente el amplio rango de influencia de estas moléculas, que afecta directa o indirectamente a casi todas las vías celulares. El principal factor en el impacto de los microARN es la regulación de la expresión genética mediante la represión, el bloqueo de su traducción o la destrucción de transcripciones específicas de ARN mensajero (ARNm).
La actividad reguladora de los microARN está estrechamente relacionada con el complejo proteico RISC, que es responsable de dos etapas importantes: el reconocimiento del objetivo y la implementación del efecto regulador. El reconocimiento del ARNm objetivo y la unión están garantizados por la interacción complementaria del sitio de reconocimiento del microARN y la región de reconocimiento (elemento de respuesta de miARN, MRE) ubicada en el ARNm objetivo. Los efectos reguladores de los microARN se obtienen gracias a muchos componentes proteicos de RISC, como AGO (Argonaute), que, después del reconocimiento y la unión al objetivo, desencadena el proceso de desarrollo del efecto en el objetivo.
El complejo proteico RISC es una estructura muy importante que participa en muchos procesos que regulan la actividad genética (Figura 2A).
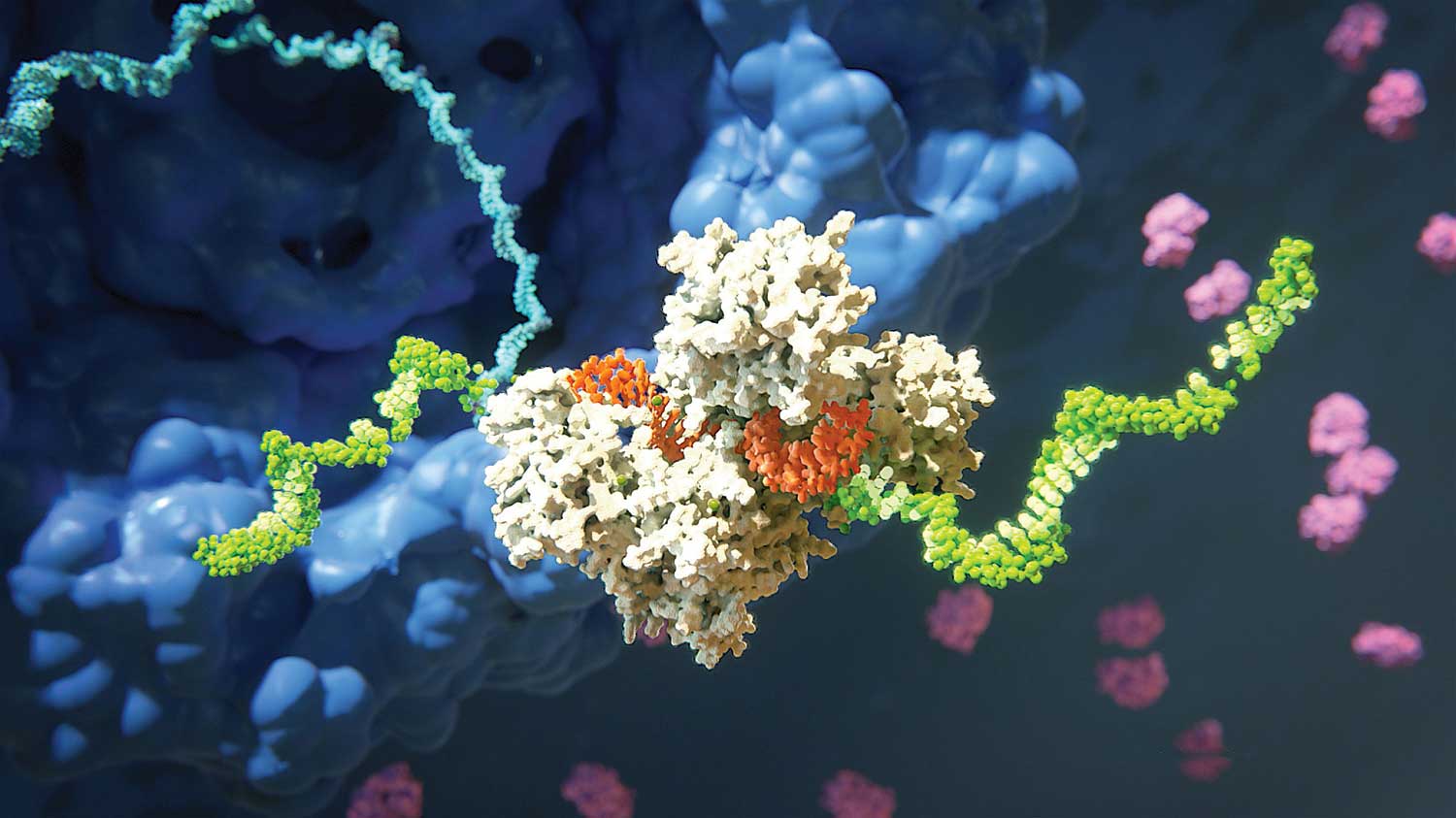
Figura 2A. Complejo inducido por ARN (RISC) que escinde el ARNm diana. RISC (que se muestra aquí en blanco) incluye la proteína Argonauta y una molécula de ARN monocatenario (naranja) derivada de un pequeño ARN de interferencia (ARNip). Se produce la escisión del ARNm (verde) que es complementario a una de las cadenas de microARN. Este proceso finaliza con la degradación del ARNm diana.
La interacción del microARN con el complejo proteico RISC se produce según el siguiente escenario: una de las hebras del dúplex de miARN (principal) se inserta en el complejo proteico, formando una estructura que se une al extremo 3′ no traducido (3′ UTR) de el ARNm objetivo. Después de esto, el microARN actúa sobre su objetivo, lo que puede provocar la destrucción del ARNm por las nucleasas o la supresión de su traducción (Figura 2B).
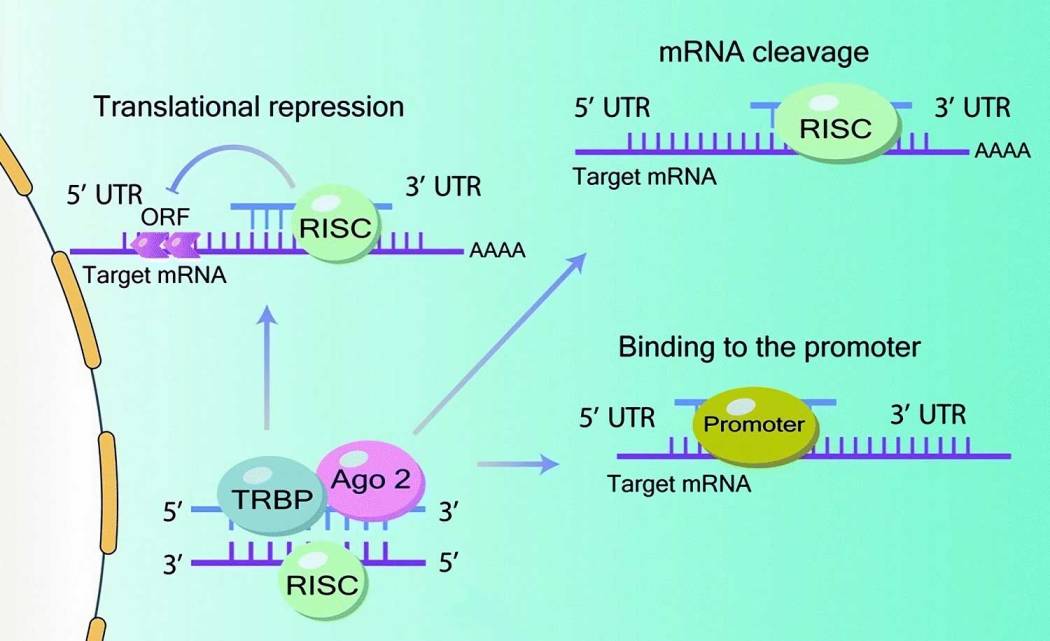
Figura 2B. Mecanismo de acción del microARN: después de la exportación del pre-microARN desde el núcleo (utilizando exportina-5 dependiente de Ran-GTP) al citoplasma y su conversión en microARN maduro, el ensamblaje del complejo RISC se produce mediante la interacción del ARN- unión de la proteína TAR (TRBP) y Argonaute 2 (Ago 2) para apuntar a las regiones 3′ no traducidas (3′-UTR) de los ARNm objetivo. Luego, el proceso de traducción se inhibe mediante la unión complementaria al extremo 3′ no traducido (3′ UTR) del ARNm objetivo. Sin embargo, también son posibles otro tipo de interacciones: degradación del ARNm o inhibición de la traducción por unión al promotor de la región codificante y 5’UTR.
En general, se cree que los miARN animales funcionan abrumadoramente mediante la represión traduccional, mientras que los miARN vegetales funcionan mediante la represión postranscripcional de la expresión génica. Al mismo tiempo, los microARN pueden influir en la traducción en las etapas de iniciación y elongación. Curiosamente, además de la supresión transcripcional, se han descubierto otros mecanismos de acción de los microARN. Por lo tanto, los microARN pueden estabilizar los ARNm y mejorar su traducción, actuar como un “cebo”, evitando la interacción de factores proteicos con sus objetivos de ARN y también participar en la maduración de otros microARN.