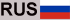




Нобелевскую премию по физиологии и медицине получили американцы Виктор Эмброс из Массачусетского университета и Гэри Равкан из Гарвардской медицинской школы «за открытие микроРНК и ее роли в посттранскрипционной регуляции генов».
Информацию, хранящуюся в хромосомах, можно сравнить с инструкцией для всех клеток организма. Каждая клетка содержит один и тот же набор генов и одинаковый набор инструкций. Тем не менее разные типы клеток, в частности мышечные и нервные, довольно сильно отличаются друг от друга и выполняют разные функции. Эти различия обеспечивает многоуровневая регуляция генов, благодаря которой в клетках каждого типа активен свой набор генов и действуют нужные инструкции. Лауреаты открыли один из механизмов генной регуляции, где ключевую роль играют очень короткие молекулы РНК, названные микроРНК. Чем сложнее организм, тем больше в нем разновидностей микроРНК: например, у человека их обнаружено уже около 2000. Открытие лауреатов еще не используется в медицине, но, возможно, окажется полезным в лечении онкологических заболеваний.
Хотя микроРНК и закодированы в небольшой части генома, в нормальных физиологических условиях они выполняют ряд ключевых функций в развитии, дифференцировке клеток, регуляции экспрессии генов, клеточном цикле и апоптозе, пишет журнал Биомолекула.
Эти маленькие регуляторы — крайне важный инструмент организма. Нарушение в работе одной или небольшого подмножества микроРНК оказывает глубокое влияние на особенности экспрессии сотен мРНК. А это уже, в свою очередь, как по цепочке, влечет за собой нарушения в протекании основополагающих биологических процессов клетки. Итогом всех этих изменений зачастую становится опухоль. Так, в одно мгновение из важных контролёров клеточного цикла микроРНК превратились в диверсантов для собственного организма. Такое поведение свойственно онкогенным микроРНК.
Гидрогелевый каркас из самоорганизующейся РНК-тройной спирали для модуляции микроРНК в микроокружении опухоли. Криосекция адгезивного гидрогеля дендример—декстран (толщина 12 мкм) с изображением морфологии адгезива (декстрановый альдегид был помечен Alexa Fluor 405). Красные пятна представляют собой наночастицы с тройной спиралью, содержащие олигонуклеотиды Q705 (красный) и Q570 (зеленый).
Но есть еще и онкосупрессорные микроРНК, которые, будто пытаясь реабилитироваться, ингибируют процессы опухолеобразования и способствуют ремиссии рака. Такое разделение весьма неоднозначно, так как некоторые микроРНК в одних условиях могут быть онкосупрессорными, а в других — онкогенными.
Исходя из настолько противоположных функций, сложно сделать однозначные выводы о роли микроРНК в процессах канцерогенеза, однако многочисленные исследования подробно описали их влияние на клеточный цикл, метастазирование и ангиогенез в контексте развития рака.
Далеко не каждая болезнь может похвастаться таким резонансом в обществе, как рак. Этот собирательный термин охватывает широкую группу заболеваний, которые без преувеличения можно назвать «глобальной проблемой». Вопрос лечения рака остается актуальным и по сей день. Согласно данным ВОЗ, в 2020 году эта болезнь унесла около 10 миллионов жизней, что соответствует примерно одной шестой от общего числа всех смертей.
МикроРНК (miRNA) представляют собой класс небольших некодирующих молекул РНК, последовательности средней длиной в 20 нуклеотидов. Они обычно собираются в ядре, следуя двум основным путям: каноническому и неканоническому.
Первый связан с транскрипцией первичной микроРНК (при-микроРНК, от англ. primary — «первичная») при помощи РНК-полимераз II или III. Эта при-микроРНК процессируется в предшественник микроРНК (пре-микроРНК) рибонуклеазой по имени Дроша (Drosha). В конечном результате экспортируется из ядра в цитозоль, где эндонуклеаза семейства РНКаз III (Dicer) преобразует пре-миРНК в дуплекс микроРНК (рисунок 1).
Созревание микроРНК завершается образованием рибонуклеопротеинового комплекса — miRISC (комплекс молчания, индуцируемого микроРНК). В цитоплазме происходит ассоциация микроРНК с белками TRBP (transactivation response RNA-binding protein), PACT (protein kinase R activating protein) и одного из белков семейства Argonaute, в результате чего и образуется комплекс RISC. Только после образования этого комплекса микроРНК приобретает функциональную активность. Другая же цепь микроРНК (пассажирская) часто (но не всегда) удаляется.
По описанному механизму происходит биогенез большей части микроРНК, но этот путь не единственный. Около 40% микроРНК животных кодируются в интронах белок-кодирующих генов и, следовательно, многие транскрипты при-микроРНК являются одновременно пре-мРНК и при-микроРНК (Cai et al., 2004; Kim et al., 2009). Такие интроны, называемые миртронами, сплайсируются и формируют шпильки микроРНК, подходящие для расщепления Dicer. В таком случае микроРНК минует некоторые этапы канонического пути.
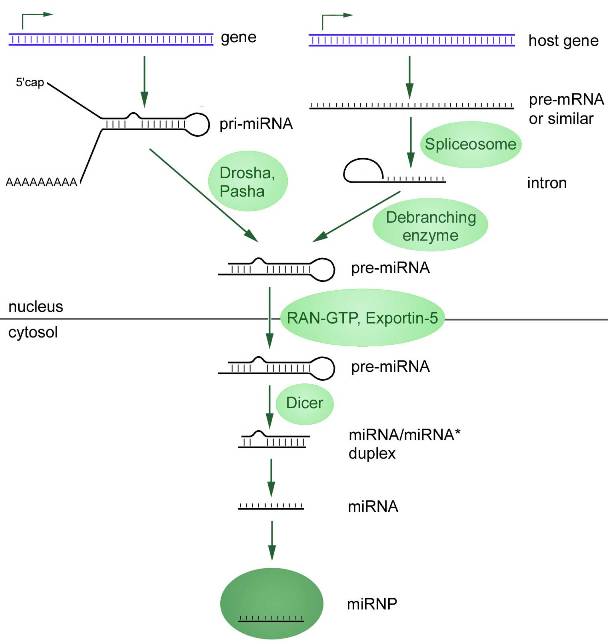
Рисунок 1. Два пути биогенеза микроРНК, начинающиеся в клеточном ядре с транскрипции. Канонический путь представлен слева: при-микроРНК подвергается 5’- кэпированию и часто 3’-полиаденилированию (исключение составляют гены, прилегающие к Alu-повторам). C помощью ферментов Drosha Pasha при-микроРНК обрезается, преобразуясь в миРНК-предшественницу (пре-микроРНК) с более короткой последовательностью. Неканонический путь: из интронов (миртронов) транскрибируется первичная микроРНК (при-микроРНК) в ядре, которая дальше сплайсируется сплайсосомами, а после формирует характерную шпильку. Образовавшаяся по итогам двух путей синтеза пре-микроРНК экспортируется из ядра в цитоплазму, где и преобразуется в зрелую микроРНК посредствам фермента Dicer.
Существует куда большее количество всевозможных вариантов биогенеза микроРНК, но эти два можно назвать основными.
Не стоит недооценивать роль микроРНК. Это очень важные регуляторы многих биологических процессов. Тот факт, что примерно треть генов, кодирующих белки, контролируется микроРНК, явно указывает на широкий круг влияния этих молекул, в который прямо или косвенно попадают почти все клеточные пути. Основной фактор воздействия микроРНК — регулирование экспрессии генов посредством репрессии, блокировки их трансляции или же разрушения специфических транскриптов матричной РНК (мРНК).
Регуляторная деятельность микроРНК тесно связана с белковым комплексом RISC, который отвечает за два важных этапа: распознавание мишени и осуществление регуляторного эффекта. Распознавание мРНК-мишени и связывание обеспечиваются комплементарным взаимодействием участка узнавания микроРНК и узнаваемого участка (miRNA response element, MRE), находящегося в мРНК-мишени. Регуляторные же эффекты микроРНК обеспечиваются благодаря многим белковым составляющим RISC, таким как AGO (Argonaute), который после узнавания и связывания с мишенью и запускает процесс развития эффекта на мишень.
Белковый комплекс RISC — это очень важная структура, принимающая участие во многих процессах регуляции активности генов (рисунок 2А).
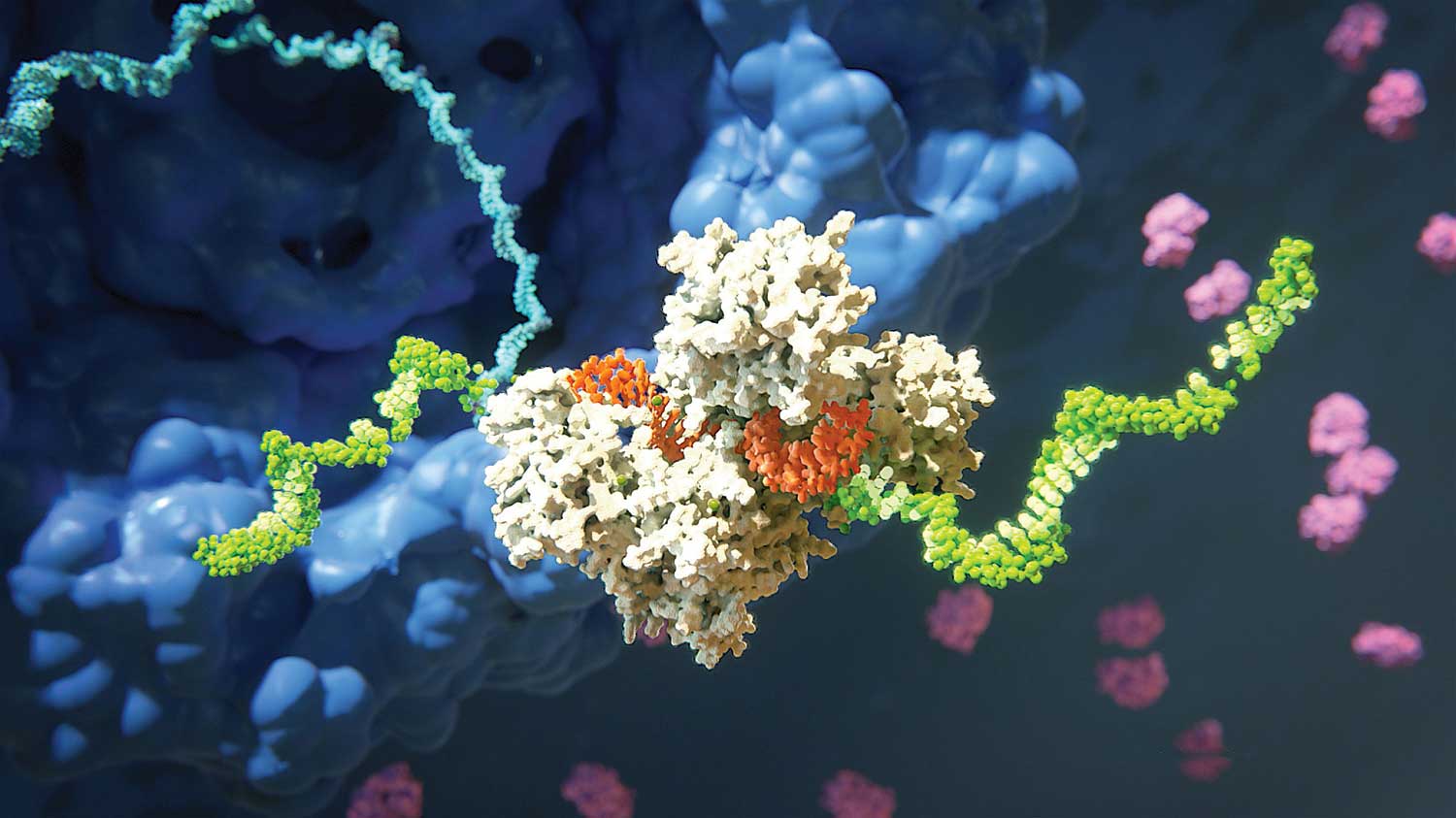
Рисунок 2А. РНК-индуцированный комплекс (RISC), расщепляющий мРНК-мишень. RISC (показан здесь белым цветом) включает в себя белок Argonaute и одноцепочечную молекулу РНК (оранжевый), полученную из малой интерферирующей РНК (миРНК). Происходит расщепление мРНК (зеленой), комплементарной одной из цепей микроРНК. Завершается данный процесс деградацией мРНК-мишени.
Взаимодействие микроРНК с белковым комплексом RISC происходит по следующему сценарию: одна из цепей дуплекса микроРНК (ведущая) внедряется в белковый комплекс, образуя структуру, связывающую 3’-нетранслируемый конец (3’ UTR) мРНК-мишени. После этого микроРНК воздействует на свою мишень, результатом чего может стать разрушение мРНК нуклеазами или же подавление её трансляции (рисунок 2Б).
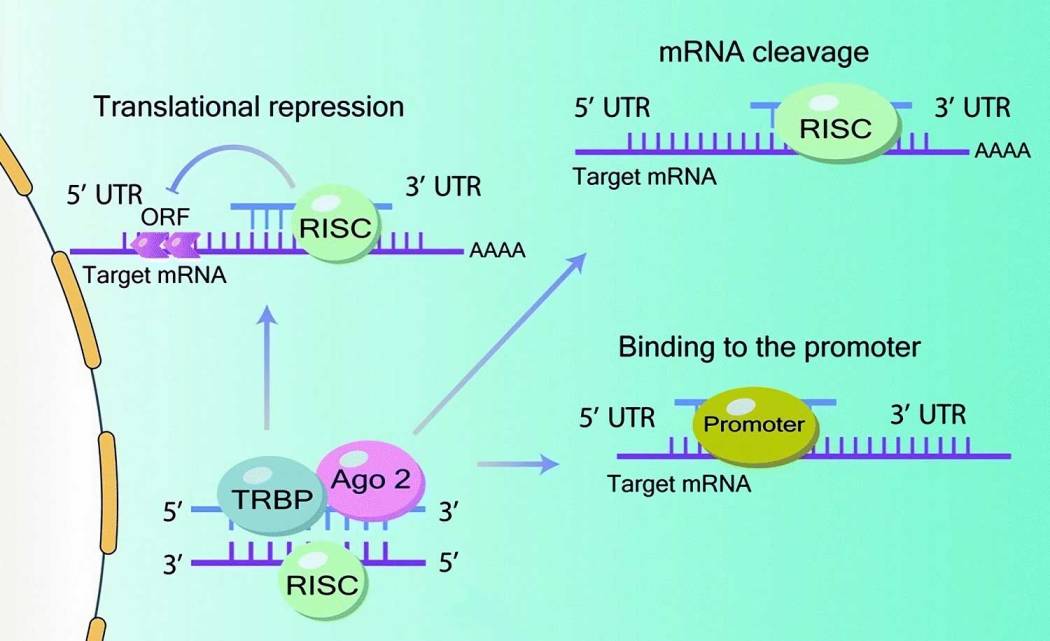
Рисунок 2Б. Механизм действия микроРНК: после экспорта пре-микроРНК из ядра (при помощи Ran-GTP-зависимого экспортина-5 ) в цитоплазму и преобразования до зрелой микроРНК происходит сборка RISC-комплекса при взаимодействии РНК-связывающего белка TAR (TRBP) и Argonaute 2 (Ago 2) для нацеливания на 3’-нетранслируемые области (3’-UTR) целевых мРНК. Затем процесс трансляции ингибируется путем комплементарного связывания с 3’-нетранслируемым концом (3’ UTR) целевой мРНК. Однако возможны и другие варианты взаимодействий: деградация мРНК или же ингибирование трансляции вследствие связывания с промотором кодирующей области и 5′ UTR.
В целом считается, что микроРНК животных в подавляющем большинстве функционируют благодаря репрессии трансляции, тогда как микроРНК растений функционируют посредством посттранскрипционного подавления экспрессии генов. При этом микроРНК способны влиять на трансляцию на этапах инициации и элонгации. Интересно, что, помимо подавления транскрипции, обнаружены и другие механизмы действия микроРНК. Так, микроРНК могут стабилизировать мРНК и усиливать их трансляцию, выполнять функцию «приманки», препятствуя взаимодействию белковых факторов с их РНК-мишенями, а также участвовать в процессе созревания других микроРНК.