



Le prix Nobel de physiologie ou médecine a été décerné aux Américains Victor Ambrose de l’Université du Massachusetts et Gary Ravkan de la Harvard Medical School «pour la découverte du microARN et son rôle dans la régulation post-transcriptionnelle des gènes».
Les informations stockées dans les chromosomes peuvent être comparées aux instructions destinées à toutes les cellules du corps. Chaque cellule contient le même ensemble de gènes et le même ensemble d’instructions. Cependant, différents types de cellules, en particulier les cellules musculaires et nerveuses, sont très différentes les unes des autres et remplissent des fonctions différentes. Ces différences sont assurées par une régulation génétique à plusieurs niveaux, grâce à laquelle dans chaque type de cellule un ensemble différent de gènes est actif et les instructions nécessaires sont appliquées. Les lauréats ont découvert l’un des mécanismes de régulation des gènes, dans lequel des molécules d’ARN très courtes appelées microARN jouent un rôle clé. Plus l’organisme est complexe, plus il contient de variétés de microARN : par exemple, environ 2000 d’entre eux ont déjà été découverts chez l’homme. La découverte des lauréats n’a pas encore été utilisée en médecine, mais elle pourrait être utile dans le traitement du cancer. .
Bien que les microARN soient codés dans une petite partie du génome, dans des conditions physiologiques normales, ils remplissent un certain nombre de fonctions clés dans le développement, la différenciation cellulaire, la régulation de l’expression des gènes, le cycle cellulaire et l’apoptose, écrit la revue Biomolecule.
Ces petits régulateurs sont un outil extrêmement important pour le corps. La perturbation d’un ou d’un petit sous-ensemble de microARN a des effets profonds sur les modèles d’expression de centaines d’ARNm. Et ceci, à son tour, comme dans une chaîne, entraîne des perturbations au cours des processus biologiques fondamentaux de la cellule. Le résultat de tous ces changements est souvent une tumeur. Ainsi, en un instant, les microARN sont passés d’importants contrôleurs du cycle cellulaire à des saboteurs de leur propre corps. Ce comportement est typique des microARN oncogènes.
Échafaudage d’hydrogel à triple hélice d’ARN auto-assemblé pour la modulation des microARN dans le microenvironnement tumoral. Cryosection d’un hydrogel adhésif dendrimère-dextrane (12 µm d’épaisseur) montrant la morphologie de l’adhésif (l’aldéhyde de dextrane a été marqué avec Alexa Fluor 405). Les points rouges représentent des nanoparticules à triple hélice contenant les oligonucléotides Q705 (rouge) et Q570 (vert).
Mais il existe également des microARN suppresseurs de tumeurs qui, comme s’ils tentaient de se réhabiliter, inhibent les processus de formation de tumeurs et favorisent la rémission du cancer. Cette division est très ambiguë, puisque certains microARN peuvent être suppresseurs de tumeurs dans certaines conditions et oncogènes dans d’autres.
Sur la base de fonctions aussi opposées, il est difficile de tirer des conclusions claires sur le rôle des microARN dans la cancérogenèse, mais de nombreuses études ont détaillé leur influence sur le cycle cellulaire, les métastases et l’angiogenèse dans le contexte du développement du cancer.
Toutes les maladies ne peuvent pas se vanter d’avoir une telle résonance dans la société que le cancer. Ce terme collectif couvre un large groupe de maladies qui, sans exagération, peuvent être qualifiées de « problème mondial ». La question du traitement du cancer reste d’actualité à ce jour. Selon l’OMS, la maladie a fait environ 10 millions de morts en 2020, soit environ un sixième de tous les décès.
Les microARN (miARN) sont une classe de petites molécules d’ARN non codantes, des séquences d’une longueur moyenne de 20 nucléotides. Ils sont généralement assemblés dans le noyau, en suivant deux chemins principaux : canonique et non canonique.
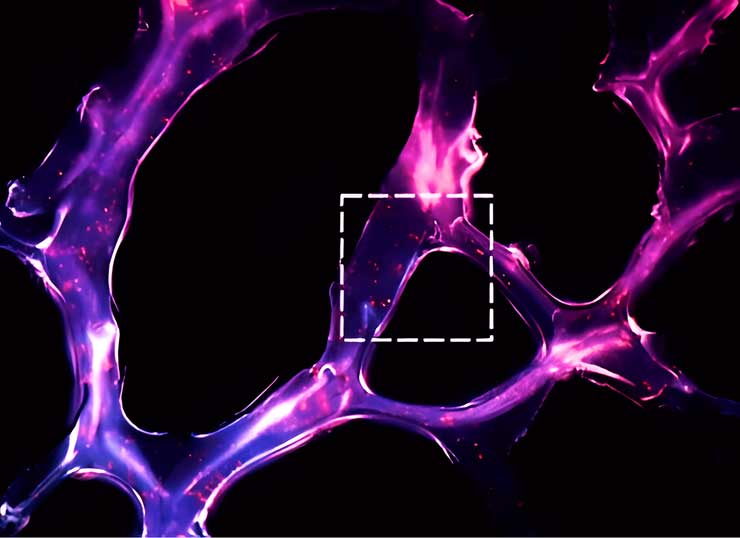
Le premier est associé à la transcription du microARN primaire (pri-microARN, de l’anglais primaire – «primaire») à l’aide des ARN polymérases II ou III. Ce pri-miARN est transformé en précurseur de microARN (pré-miARN) par une ribonucléase appelée Drosha. Le résultat final est exporté du noyau vers le cytosol, où une endonucléase de la famille RNase III (Dicer) convertit le pré-miARN en un duplex de miARN (Figure 1).
La maturation des microARN se termine par la formation d’un complexe ribonucléoprotéique, miRISC (microRNA-induced silencing complexe). Dans le cytoplasme, les microARN s’associent aux protéines TRBP (protéine de liaison à l’ARN de réponse de transactivation), PACT (protéine activatrice de la protéine kinase R) et à l’une des protéines de la famille Argonaute, à la suite desquelles se forme le complexe RISC. Ce n’est qu’après la formation de ce complexe que les microARN acquièrent une activité fonctionnelle. L’autre brin de microARN (passager) est souvent (mais pas toujours) supprimé.
Selon le mécanisme décrit, la biogenèse de la plupart des microARN se produit, mais cette voie n’est pas la seule. Environ 40% des miARN animaux sont codés dans des introns de gènes codant pour des protéines et, par conséquent, de nombreux transcrits de pri-miARN sont à la fois des pré-ARNm et des pri-miARN (Cai et al., 2004; Kim et al., 2009). De tels introns, appelés mirtrons, sont épissés pour former des épingles à cheveux miARN adaptées au clivage Dicer. Dans ce cas, le microARN contourne certaines étapes de la voie canonique.
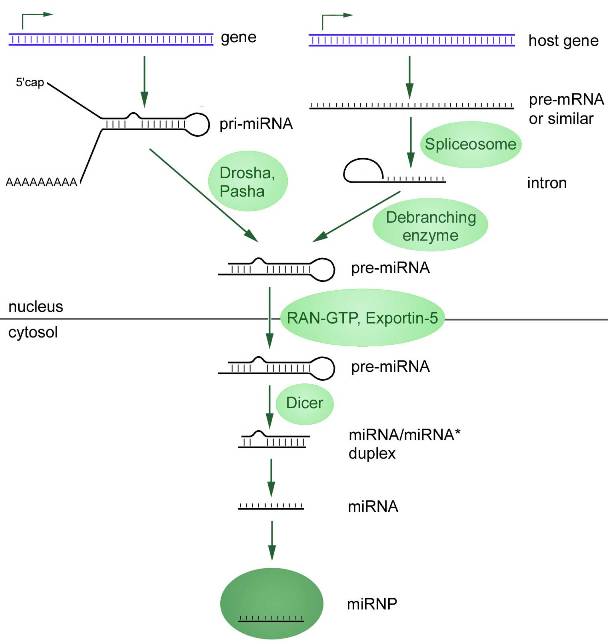
Figure 1. Deux voies de biogenèse des microARN, commençant dans le noyau cellulaire avec transcription. La voie canonique est présentée à gauche : le pri-microARN subit un coiffage en 5′ et souvent une polyadénylation en 3′ (à l’exception des gènes adjacents aux répétitions Alu). À l’aide des enzymes Drosha Pasha, le pri-microARN est coupé, se transformant en un miARN précurseur (pré-microARN) avec une séquence plus courte. Voie non canonique : le microARN primaire (pri-microARN) est transcrit à partir des introns (mirtrons) dans le noyau, qui est ensuite épissé par des spliceosomes, puis forme une épingle à cheveux caractéristique. Le pré-microARN formé à la suite de deux voies de synthèse est exporté du noyau vers le cytoplasme, où il est converti en microARN mature grâce à l’enzyme Dicer.
Il existe de nombreuses autres variantes différentes de la biogenèse des microARN, mais ces deux-là peuvent être qualifiées de principales.
Le rôle des microARN ne doit pas être sous-estimé. Ce sont des régulateurs très importants de nombreux processus biologiques. Le fait qu’environ un tiers des gènes codant pour les protéines soient contrôlés par des microARN indique clairement le large éventail d’influences de ces molécules, qui affectent directement ou indirectement presque toutes les voies cellulaires. Le principal facteur d’impact des microARN est la régulation de l’expression des gènes par la répression, le blocage de leur traduction ou la destruction de transcrits spécifiques d’ARN messager (ARNm).
L’activité régulatrice des microARN est étroitement liée au complexe protéique RISC, qui est responsable de deux étapes importantes: la reconnaissance de la cible et la mise en œuvre de l’effet régulateur. La reconnaissance de l’ARNm cible et la liaison sont assurées par l’interaction complémentaire du site de reconnaissance du microARN et de la région de reconnaissance (élément de réponse miARN, MRE) située dans l’ARNm cible. Les effets régulateurs des microARN sont assurés grâce à de nombreux composants protéiques de RISC, comme l’AGO (Argonaute), qui, après reconnaissance et liaison à la cible, déclenche le processus de développement de l’effet sur la cible.
Le complexe protéique RISC est une structure très importante impliquée dans de nombreux processus régulant l’activité des gènes (Figure 2A).
Figure 2A. Complexe induit par l’ARN (RISC) qui clive l’ARNm cible. RISC (représenté ici en blanc) comprend la protéine Argonaute et une molécule d’ARN simple brin (orange) dérivée de petits ARN interférents (siARN). Le clivage de l’ARNm (vert) complémentaire d’une des chaînes de microARN se produit. Ce processus se termine par la dégradation de l’ARNm cible.
L’interaction du microARN avec le complexe protéique RISC se produit selon le scénario suivant : l’un des brins du duplex de miARN (principal) est inséré dans le complexe protéique, formant une structure qui lie l’extrémité 3′ non traduite (3′ UTR) de l’ARNm cible. Le microARN agit ensuite sur sa cible, ce qui peut entraîner la destruction de l’ARNm par des nucléases ou la suppression de sa traduction (Figure 2B).
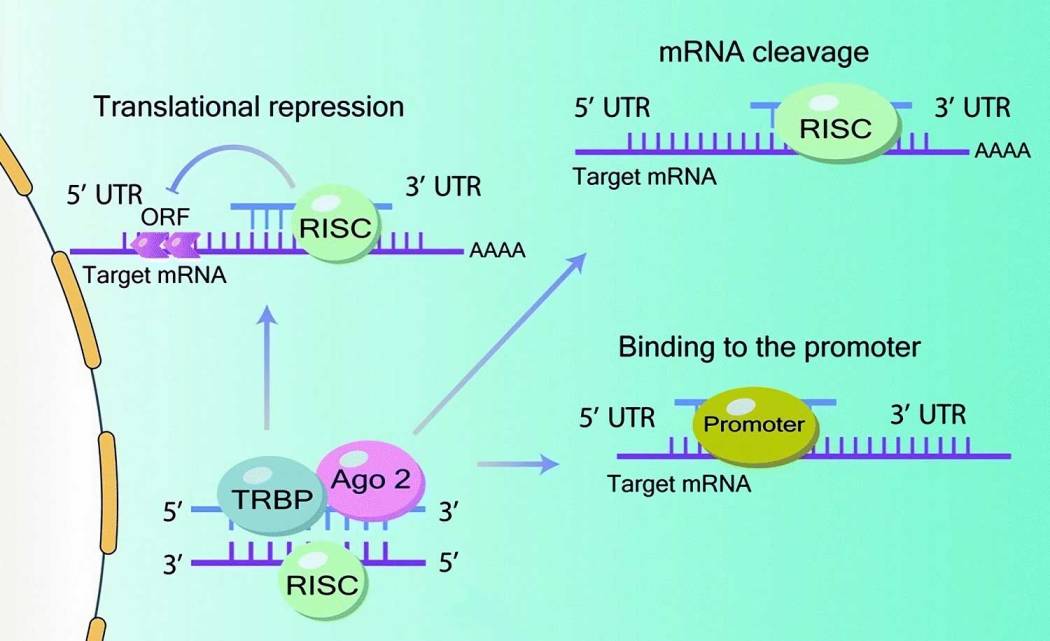
Figure 2B. Mécanisme d’action du microARN : après l’exportation du pré-microARN du noyau (en utilisant l’exportine-5 dépendante de Ran-GTP) vers le cytoplasme et la conversion en microARN mature, l’assemblage du complexe RISC se produit grâce à l’interaction de l’ARN- protéine de liaison TAR (TRBP) et Argonaute 2 (Ago 2) pour cibler les régions 3′ non traduites (3′-UTR) des ARNm cibles. Le processus de traduction est ensuite inhibé par une liaison complémentaire à l’extrémité 3′ non traduite (3′ UTR) de l’ARNm cible. Cependant, d’autres types d’interactions sont également possibles : dégradation de l’ARNm ou inhibition de la traduction due à la liaison au promoteur de la région codante et à la 5′ UTR.
En général, on pense que les miARN animaux fonctionnent essentiellement par répression traductionnelle, tandis que les miARN végétaux fonctionnent par répression post-transcriptionnelle de l’expression des gènes. Dans le même temps, les microARN sont capables d’influencer la traduction aux étapes d’initiation et d’élongation. Fait intéressant, outre la suppression transcriptionnelle, d’autres mécanismes d’action des microARN ont été découverts. Ainsi, les microARN peuvent stabiliser les ARNm et améliorer leur traduction, agir comme un « appât », empêchant l’interaction des facteurs protéiques avec leurs cibles ARN, et également participer à la maturation d’autres microARN.

